SPLICING
Karakteristik mRNA hasil transkripsi pada jasad eukariotik yakni berupa pre-mRNA (transkrip primer, primary transcript) yang meupakan sekuens yang tidak diterjemahkan atau disebut dengan intron dan adanya sekuens yang diterjemahkan yang disebut dengan ekson (Kim et al., 2005). Jumlah ekson biasanya lebih banyak dari pada jumlah intron. Jumlah intron dalan suatu gen sangat bervariasi, meskipun jumlah ekson lebih banyak daripada intron, namun sekuens intron lebih panjang. Peristiwa pemotongan segmen sekuens intron dan pembentukan mRNA matang (mature) dari penggabungan dari berbagai sekuens ekson disebut sebagai peristiwa RNA splicing (Freifelder, 1987). Adapun mekanisme singkat splicing disajikan dalam Gambar 1.

Mekanisme Splicing
Mekanisme splicing membutuhkan sekuens nukleotida yang spesifik pada daerah dua ujung intron, yakni pada ujung 5′ terdapat sekuens 5′–GU–3′ dan pada ujung 3′ terdapat sekuens 5′–AG–3′. Ujung sisi 3′ bertujuan untuk mengenali daerah downstream splicing, sementara ujung sisi 5′ bertujuan untuk mengenali daerah upstream splicing (Clark, 2010; Lasda et al.,2010; Turner et al., 1997). Pada Gambar 2 ditunjukkan pola sekuens spesifik pada b-globin manusia yang mana garis warna hijau menunjukkan daerah intron dengan ciri khas adanya sekuens GT (GU) pada ujung 5′ dan adanya sekuens AG pada ujung 3′. Sementara warna merah adalah daerah ekson yang mana ekson 1 adalah leader sequence; ekson 2 adalah coding area; dan ekson 3 adalah untranslated sequence (Alberts et al., 2008).

Selain adanya sinyal pada ujung 5′ dan 3′, juga ditemukan adanya sekuens sinyal yang terletak di tengah intron yang yang disebut dengan branchpoint sequence yang sekuensnya berupa 5′–CURAY–3′. Pada verterbrata, R = purin, Y = pirimidin, namun pada khamir ditemukan sekuens spesifik yakni 5′–UACUAAC–3′ (Lasda et al.,2010; Turner et al., 1997).
Sinyal-sinyal pada ujung 5′, 3′, dan branchpoint sequence akan displicing oleh snRNA (small nuclear RNA) yang akan berasosiasi dengan suatu protein membentuk kompleks protein small ribonuclear proteins (snRNPs, dibaca “snurp”) yang terdiri dari U1,U2, U4, U5, dan U6 (Lasda et al., 2010; Yuwono, 2007). Snurp U1 akan mengenali dan mengikat ujung 5′ dari intron; snurp U2 akan mengikat branchpoint sequence; snurp U2AF (U2 accesore factor) akan mengikat ujung 3′; dan snurp U4/U6 akan mengikat snurp U2 dan U6 (Gambar 3). Kompleks antara snRNPs dengan pre-mRNA akan membentuk suatu kompleks yang dinamakan spilceosome (Sanford&Caceres, 2004). Spliceosome tersebut akan membentuk gulungan (loop) pada intron dan selanjutnya intron dipotong dari pre-mRNA dalam bentuk lariat atau seperti tali laso seperti yang ditunjukkan pada Gambar 4 (Aitken et al., 2011; Clark, 2010, Turner et al., 1997).

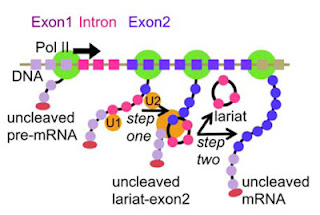
Gambar 4. Splicing dapat terjadi selama elongasi transkripsi. RNA polimerase II (ditampilkan dalam warna hijau) mentranskripsi Exon1, Intron dan Exon2 (warna RNA menunjukkan wilayah yang sesuai dari DNA, dan warna oval merah menunjukkan cap pada ujung 5′ dari RNA). Splicing dapat terjadi setelah mentranskripsi urutan tertentu yang memicu perakitan dan aktivasi spliceosome pada RNA menmbentuk struktur lariat-ekson2 yang selanjutnya lariat akan memisah dari ekson (Aitken et al., 2011).
Pada Gambar 4 tersebut juga dijelaskan mekanisme dua langkah reaksi (two-step reaction). Langkah pertama adalah pembentukan spliceosome dengan bentuk lariat-ekson2 yang dibantu oleh RNA Polimerase II, kemudian langkah kedua adalah sisi dari ujung 3′ (3′SS) mengalami pelekatan dan selanjutnya terbentuklah mRNA yang sudah mengalami maturasi (Aitken et al, 2011; Liu et al., 2008).